Anabolic/androgenic steroids (AAS), when introduced into the body, do not necessarily remain unchanged. Enzymatic processes often convert them to different molecules. This is true of the prohormones as well: androstenedione will not necessarily remain as androstenedione, but some of it will be converted to testosterone. Although that example is well known, many other enzymatic conversions of steroids are less well-known in bodybuilding. We will be looking at these conversions in this article.
Esterases
Anabolic steroids given by intramuscular injection are usually (not always) esterified versions of the parent drugs (Figure 1). This esterification improves the oil solubility of the steroid, and reduces water solubility: the result is that the esterified drug remains stored mostly dissolved in fat, and the parent drug is released to the bloodstream only slowly, over time.
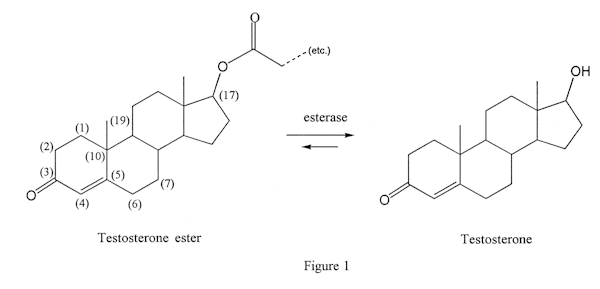
The enzymes that accomplish this transformation are called esterases, and are widely present in the body. Aside from allowing use of esterified drugs, they have another effect: they can also work in the reverse direction. Instead of removing an ester from the esterified drug, they can add an ester to the parent drug. This can increase the lifetime of the drug in the body. Thus, it could be expected that some small amount of a norandro prohormone would be converted to an esterified form, and would remain (in trace amounts) in the system for quite some time. Unfortunately, analytical techniques today are sufficient to detect nandrolone or its metabolites in extremely low concentrations. Thus, the norandro products cannot be recommended to anyone who might be subject to drug testing for AAS.
Esterases cannot work well on steroid esters that are 17alpha alkylated, because the alkylation blocks the approach of the enzyme to the ester. This is the reason that no such drugs are sold – there are no esters of Dianabol, methyltestosterone, Anadrol®, oxandrolone, Winstrol, etc., and there never will be (Figure 2). They would be inactive.
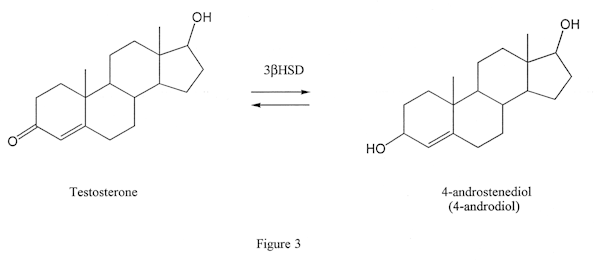
3beta hydroxysteroid dehydrogenase (3bHSD)
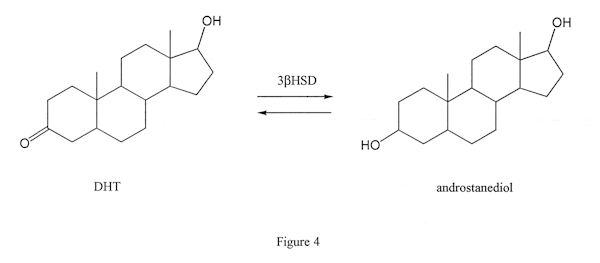
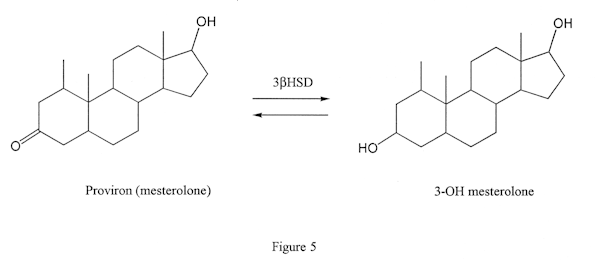
This enzyme, like the esterase enzymes, can work in two directions. It can either convert a steroid that has a keto group on position 3 of the steroid (Figure 3, the left molecule) to one with a hydroxy group in the same position (Figure 3, the molecule on the right) or vice versa. The latter action is seen when androdiol is converted to testosterone by this enzyme. The former is seen when DHT is converted to androstanediol (not androstenediol) in muscle tissue (Figure 4): this is the reason DHT is not an effective anabolic in muscle tissue.) Proviron also undergoes this transformation and is deactivated in muscle tissue (Figure 5).
The conversion of DHT to androstanediol also occurs in scalp tissue, and androstanediol may be of relevance in the development of male pattern baldness.
Because the enzyme works in two directions, it cannot convert a keto steroid entirely to the hydroxy, or vice versa, because some of what is converted will then be acted upon again and return to its original state. Thus, there will always be a mixture of the two compounds. In some cases, the mixture might favor one side of the balance.
3bHSD is widely distributed in the body.
17beta-hydroxysteroid dehydrogenase (17bHSD)
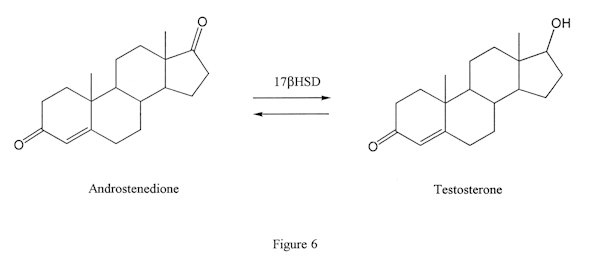
Again, this is an enzyme that can work in two directions. It can convert a steroid that has a keto group in the 17 position (Figure 6, molecule on left) to one that has a hydroxy group in the same position (Figure 6, molecule on right), or vice versa. The former conversion takes place in the case of androstenedione being converted to testosterone.
Aromatase
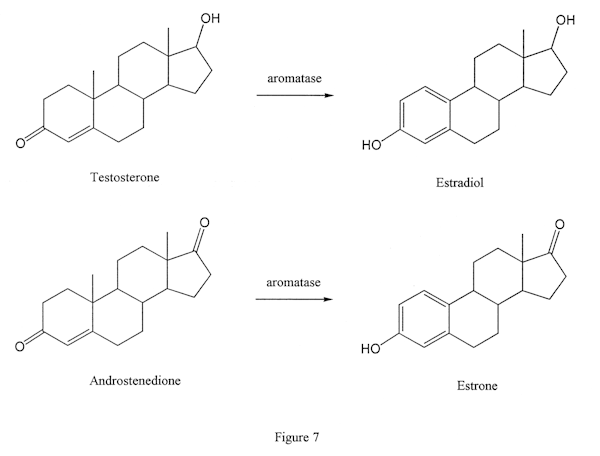
This enzyme removes the 19 methyl from AAS and aromatizes the A ring (Figure 7). This means that three, alternating double bonds are formed in that ring. Any process which produces such a pattern of bonds is called aromatization, and the enzyme is called aromatase because it accomplishes this. It is worth noting that aromatization does not require aromatase in some cases: for example, in the case of nandrolone. However, in those AAS which have a 19 methyl, the aromatase enzyme is required for aromatization, since a double bond cannot be formed at carbon 10 unless the 19 methyl is removed. (A carbon atom can have only four bonds, and there would be five in such a case.)
Aromatase is the first enzyme we have discussed where we are interested in reducing its activity. This may be done by either of two types (usually) of inhibitors: competitive inhibitors and mechanism-based (suicide) inhibitors.
Competitive inhibitors act by binding to the same site of the enzyme as the steroid molecule would. Arimidex, for example, does this. Whatever percentage of enzyme has the inhibitor bound to it, is inactive for as long as the inhibitor is bound. Thus, a small amount of inhibitor might inhibit only a small percentage of the enzyme molecules, whereas a larger amount would inhibit a higher percentage.
The inhibitor also must compete with the steroid for access to the binding site. Thus, if there is a very high amount of steroid, the steroid will be likely to “get there first” and the inhibitor will be less effective: unless its concentration is likewise increased. Thus, while Arimidex may do an excellent job at 1 mg/day for an AIDS patient using 250 mg/week of testosterone, it may fail to outcompete 1 gram per week of testosterone unless the dose is increased. This problem is true of Cytadren also.
The other type of aromatase inhibitor is one that actually destroys the enzyme, and is itself destroyed in the process. These inhibitors are called mechanism-based (suicide) inhibitors. An example of this type is Lentaron. It has not seen much use in bodybuilding and its efficacy is not clear.
Aromatase is works in only one direction: it cannot convert an aromatized steroid back to an unaromatized one.
Aromatase cannot work on DHT, Proviron, Winstrol, oxandrolone, Halotestin, or Primobolan, and there is no evidence that it can work on trenbolone. It is not clear if it works on Anadrol or perhaps on an Anadrol metabolite. There appears to be no scientific evidence that Anadrol aromatizes to estrogen, a fact pointed out by Patrick Arnold. It may be that it does not have estrogenic activity at all, but progestogenic activity. This also could be a potent inducer of gyno and bloating.
5alpha-reductase
This enzyme can do one thing and one thing only: convert a steroid with a double bond between carbon 4 and carbon 5 to one with a single bond between them (and also adding a hydrogen to each carbon so the total number of bonds remains correct.)
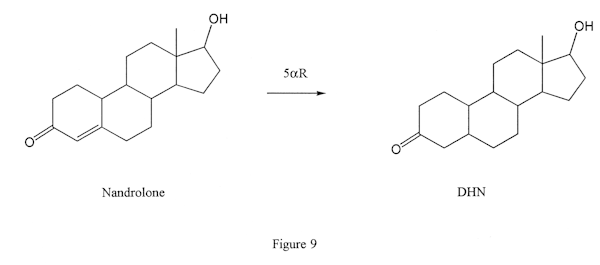
This means that it can convert testosterone to DHT (Figure 8) or nandrolone to DHN (Figure 9) but it cannot convert Winstrol or Anadrol (Figure 2) despite the uninformed claims of careless steroid authors. And although Dianabol does have a double bond between carbons 4 and 5, it nonetheless also is not converted by 5AR.
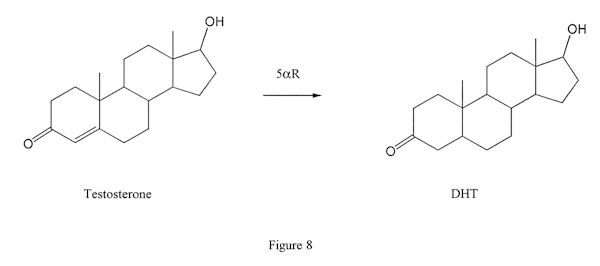
Both Type 1 and Type 2 versions (isozymes) of 5AR exist. Type 2 is present in the prostate, and Proscar is a good inhibitor of that isozyme. Type 1 is present in the scalp and skin, and unfortunately Proscar is a poor inhibitor for that type. There are a number of experimental Type 1 inhibitors, such as MK-386, which will likely be of use in treating acne or male-pattern baldness, but they are not on the market yet.
3-oxosteroid-4,5-isomerase
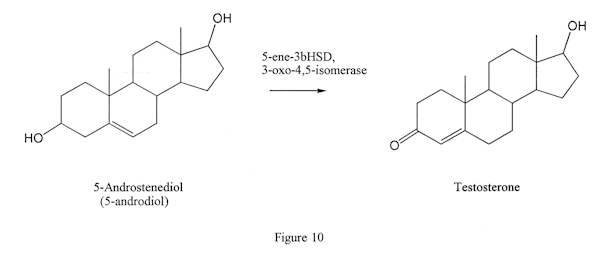
This enzyme is of no interest unless one is concerned with the conversion of DHEA or one of the 5-andro products to testosterone, where the enzyme is required to change the double bond from the 5 position to the 4 position of testosterone (Figure 10).
P450 enzymes (various)
A number of enzymes of the P450 class can hydroxylate steroids at various positions on the molecule by adding OH groups. This will deactivate the steroid, and the added hydroxy group also provides a position for further metabolism to make the steroids water soluble and more easily excreted. While some have suggested the idea of inhibiting P450 enzymes to get more “bang for the buck” from a given amount of steroid, the idea is a poor one, since there are many P450 enzymes that can act in this manner on steroids, and these enzymes are important for metabolism of many things other than steroids. Where there might be some relevance is if a drug is taken which greatly increases amount of some P450 enzymes; for example, rifampin. This would probably reduce the effectiveness of steroids by speeding their rate of metabolism.
UDP-glucuronysyltransferase (UDPGT)
This enzyme attaches what is basically a sugar molecule to an –OH group of a steroid, thus deactivating it and making it far more water soluble, and more readily excreted.
This action is reversible to a minor extent in the body by glycosidase enzymes. It is reversed to a significant extent, however, in what is called enterohepatic recycling. In this process, deactivated, glucuronidated steroids are excreted in the bile into the intestine, where bacteria then cleave the glucuronide, restoring the steroid. Much of this steroid will then be reabsorbed by the body. The result is, just because a steroid is excreted once, doesn’t mean it still won’t come back for another turn. This phenomenon is quite important in birth control, and is the reason why antibiotics can interfere with The Pill. The amount of bacteria in the intestine is reduced by the antibiotics, which reduces the degree of recycling, and thus reduces estrogen levels. It would also be the case, for an androgen user, that antibiotic use would reduce enterohepatic recycling, and thus cause a given dose of steroid to have less effect.
Men have about twice as much UDPGT activity on steroids as women do. There is also a great deal of variation between individuals.
Sulfotransferases (various types, including DHEA-ST)
These enzymes convert a –OH group of a steroid to the sulfate form, which is inactive. The process is reversible. The sulfated form provides a depot of temporarily inactivated steroid which may be later converted back to the parent steroid. This is of particular relevance with DHEA, where a large fraction of DHEA is in the sulfated form at any given moment, but is also of some relevance with the diol prohormones, which may be sulfated, and therefore inactivated, at the 3-OH position.
Sulfation generally occurs at a slower rate than glucuronidation, but nonetheless the process is still significant in the metabolism of testosterone, where conversion in the liver to the sulfate inactivates the drug and makes it more readily excreted in the bile (and again, enterohepatic recycling can occur.)
Summary
Enzymes are necessary to activate esterified steroids (the drug is inactive as long as the ester group remains present) and are necessary to deactivate steroids or to make them more water soluble and more readily excreted. They are also necessary to convert the prohormones to the active hormones. Aromatase and 5AR both convert testosterone to active compounds which may have different activities: to estrogen (in the case of aromatase) or to DHT, a more potent androgen, in the case of 5AR. 3bHSD also can convert an active compound, DHT, to androstanediol (not androstenediol) which while it is not active at the androgen receptor, may be of relevance in the development of male pattern baldness.
Control of the aromatase enzyme by inhibitors is recommended if aromatizable steroids with a C19 group (e.g., testosterone, Dianabol) are being used. A 5AR inhibitor is recommended in cases where testosterone is being used and the individual is particularly concerned about the prostate. Should a Type 1 5AR inhibitor become available, it would be of use in limiting the acne-inducing and male pattern baldness accelerating activities of testosterone.
Addendum: reading the chemical figures (this can be omitted if you are not concerned with chemistry):
Where lines meet, or a line ends with no letter at that end, there is a carbon atom. Each carbon atom will have four bonds. Lines that are drawn are bonds. In some cases, a double line may be used: this indicates a double bond that counts as two bonds. If the number of bonds is not four, then there are also hydrogen atoms present sufficient to give enough bonds. For example, \/\/\ would be a shorthand representation of:
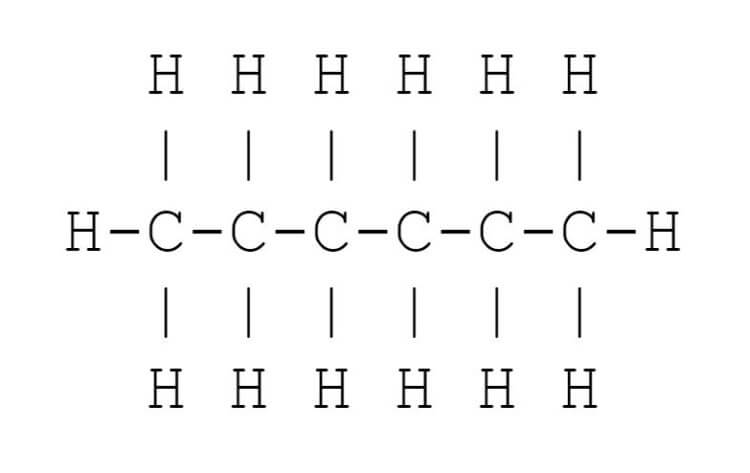
Clearly, the shorthand line form is more convenient to write, and easier for the eye to read. The steroid framework is easy to see with the line structure, but would be very cluttered if all the carbons and hydrogens were explicitly drawn.
C represents a carbon atom, H represents a hydrogen atom, O represents an oxygen atom, and N represents a nitrogen atom.
About the author
Bill Roberts is an internationally-recognized expert on anabolic steroids and performance-enhancing drugs (PEDs). He received a bachelor degree in Microbiology and Cell Science and completed the educational and research requirements for a PhD in Medicinal Chemistry at a major American university.
Bill entered the nutritional supplement industry prior to completing his doctoral thesis but his education was invaluable so far as being able to design/improve nutritional supplement compounds, since it was in the field of designing drug molecules and secondarily some work in transdermal delivery.
His education was not specifically "geared" toward anabolic steroids other than expertise with pharmacological principles having broad applications. This has allowed Bill to provide unique insight into the field of anabolic pharmacology with knowledge of points which he would not have known otherwise.
Leave a Reply
You must be logged in to post a comment.