
Super Cows and Mighty Mice
In 1997, scientists McPherron and Lee revealed to the public the ‘secret’ of an anomaly that livestock breeders have capitalized since the late 1800’s: the gene responsible for big beefy cows (1). More than a century ago, livestock breeders in Europe observed that some of their cattle were more muscled than others. Being dabblers in genetics, they selectively bred these cattle to increase the progeny displaying this trait. Thus two breeds of cattle (Belgian Blue and Piedmontese) were developed that typically exhibit an increase in muscle mass relative to other conventional cattle breeds. Little did they know that many years later Mighty Mouse would be more than merely a cartoon.
A team of scientists led by McPherron and Lee at John Hopkins University was investigating a group of proteins that regulate cell growth and differentiation. During their investigations they discovered the gene that may be responsible for the phenomenon of increased muscle mass, also called ‘double-muscling’ (1, 2). Myostatin, the protein that the gene encodes, is a member of a superfamily of related molecules called transforming growth factors beta (TGF-b ). It is also referred to as growth and differentiation factor-8 (GDF-8). By knocking out the gene for myostatin in mice, they were able to show that the transgenic mice developed two to three times more muscle than mice that contained the same gene intact. Lee commented that the myostatin gene knockout mice “look like Schwarzenegger mice.” (3).
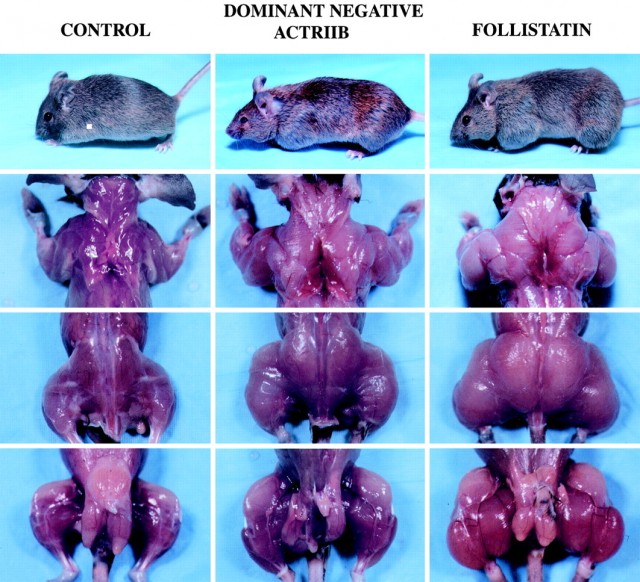
Further exploration of genes present in skeletal muscle in the two breeds of double-muscled cattle revealed mutations in the gene that codes for myostatin. The double-muscling trait of the myostatin gene knockout mice and the double-muscled cattle demonstrates that myostatin performs the same biological function in these two species. Apparently, myostatin may inhibit the growth of skeletal muscle. Knocking out the gene in transgenic mice or mutations in the gene such as in the double-muscled cattle result in larger muscle mass. This discovery has paved the way for a plethora of futuristic implications from breeding super-muscled livestock to treatment of human muscle wasting diseases.
Researchers are developing methods to interfere with expression and function of myostatin and its gene to produce commercial livestock that have more muscle mass and less fat content. Myostatin inhibitors may be developed to treat muscle wasting in human disorders such as muscular dystrophy. However, several public media sources immediately raised the issue of abusing myostatin inhibitors by athletes. In addition, a hypothesis has been put forth that a genetic propensity for high levels of myostatin is responsible for the lack of muscle gain in weight trainees. Accordingly, this article presents a look at the science of myostatin and its implications for the athletic arena.
What is Myostatin?
Growth Factors
Before we can understand the implications of tampering with myostatin and its gene, we must learn what myostatin is and what it does. Higher organisms are comprised of many different types of cells whose growth, development and function must be coordinated for the function of individual tissues and the entire organism. This is attainable by specific intercellular signals, which control tissue growth, development and function. These molecular signals elicit a cascade of events in the target cells, referred to as cell signaling, leading to an ultimate response in or by the cell.
Classical hormones are long-range signaling molecules (called endocrine). These substances are produced and secreted by cells or tissues and circulated through the blood supply and other bodily fluids to influence the activity of cells or tissues elsewhere in the body. However, growth factors are typically synthesized by cells and affect cellular function of the same cell (autocrine) or another cell nearby (paracrine). These molecules are the determinants of cell differentiation, growth, motility, gene expression, and how a group of cells function as a tissue or organ.
Growth factors (GF) are normally effective in very low concentrations and have high affinity for their corresponding receptors on target cells. For each type of GF there is a specific receptor in the cell membrane or nucleus. When bound to their ligand, the receptor-ligand complex initiates an intracellular signal inside of the cell (or nucleus) and modifies the cell’s function.
A GF may have different biological effects depending on the type of cell with which it interacts. The response of a target cell depends greatly on the receptors that cell expresses. Some GFs, such as insulin-like growth factor-I, have broad specificity and affect many classes of cells. Others act only on one cell type and elicit a specific response.
Many growth factors promote or inhibit cellular function and may be multifactoral. In other words, two or more substances may be required to induce a specific cellular response. Proliferation, growth and development of most cells require a specific combination of GFs rather than a single GF. Growth promoting substances may be counterbalanced by growth inhibiting substances (and vice versa) much like a feedback system. The point where many of these substances coincide to produce a specific response depends on other regulatory factors, such as environmental or otherwise.
Transforming Growth Factors
Some GFs stimulate cell proliferation and others inhibit it, while others may stimulate at one concentration and inhibit at another. Based on their biological function, GFs are a large set of proteins. They are usually grouped together on the basis of amino acid sequence and tertiary structure. A large group of GFs is the transforming growth factor beta (TGFb ) superfamily of which there are several subtypes. They exert multiple effects on cell function and are extensively expressed.
A common feature of TGFb s is that they are secreted by cells in an inactive complex form. Consequently, they have little or no biological activity until the latent complex is broken down. The exact mechanism(s) involved in activating these latent complexes is not completely understood, but it may involve specific enzymes. This further exemplifies how growth factors are involved in a complex system of interaction.
Another common feature of TGFb s is that their biological activity is often exhibited in the presence of other growth factors. Hence, we can see that the bioactivity of TGFb s is complex, as they are dependent upon the physiological state of the target cell and the presence of other growth factors.
Myostatin
There are several TGFb s subtypes which are based on their related structure. One such member is called growth and differentiation factors (GDF) and specifically regulates growth and differentiation. GDF-8, also called myostatin, is the skeletal muscle protein associated with the double muscling in mice and cattle.
McPherron et al detected myostatin expression in later stages of development of mouse embryos and in a number of developing skeletal muscles (1). Myostatin was also detected in adult animals. Although myostatin mRNA was almost exclusively detected in skeletal muscle, lower concentrations were also found in adipose tissue.
To determine the biological role of myostatin in skeletal muscle, McPherron and associates disrupted the gene that encodes myostatin protein in rats, leading to a loss its function. The resulting transgenic animals had a gene that was rendered non-functional for producing myostatin. The breeding of these transgenic mice resulted in offspring that were either homozygous for both mutated genes (i.e. carried both mutated genes), homozygous for both wild-type genes (i.e. carried both genes with normal function) or heterozygous and carrying one mutated and one normal gene. The main difference in resulting phenotypes manifested in muscle mass. Otherwise, they were apparently healthy. They all grew to adulthood and were fertile.
Homozygous mutant mice (often called gene knockout mice) were 30% larger than their heterozygous and wild-type (normal) littermates irregardless of sex and age. Adult mutant mice had abnormal body shapes with very large hips and shoulders and the fat content was similar to the wild-type counterparts. Individual muscles from mutant mice weighed 2-3 times more than those from wild-type mice. Histological analysis revealed that increased muscle mass in the mutant mice was resultant of both hyperplasia (increased number of muscle fibers) and hypertrophy (increased size of individual muscle fibers).
Since this discovery, McPherron and other researchers investigated the presence of myostatin and possible gene mutations in other animal species. Scientists have reported the sequences for myostatin in 9 other vertebrate animals, including pigs, chickens and humans (2, 4). Research teams separately discovered two independent mutations of the myostatin gene in two breeds of double-muscled cattle: the Belgian Blue and Piedmontese (2, 5). A deletion in the myostatin gene of the Belgian Blue eliminates the entire active region of the molecule and is non-functional; and this mutation causes hypertrophy and increased muscle mass. The Piedmontese coding sequence for myostatin contains a missense mutation. That is, a point in the sequence encodes for a different amino acid. This mutation likely leads to a complete or nearly completes loss of myostatin function.
McPherron et al analyzed DNA from other purebred cattle (16 breeds) normally not considered as double-muscled and found only one similar mutation in the myostatin gene (2). The mutation was detected in one allele a single animal which was non-double-muscled. Other mutations were detected but these did not affect protein function.
Earlier studies reported high levels of myostatin in developing cattle and rodent skeletal muscles (2, 7). Furthermore, mRNA expression varied in individual muscles. Consequently, it was thought that myostatin was relegated to skeletal muscle and that the gene’s role was restricted to the development of skeletal muscle. However, A New Zealand team of researchers recently reported the detection of myostatin mRNA and protein in cardiac muscle (8).
TGF-b superfamily members are found in a wide variety of cell types, including developing and adult heart muscle cells. Three known isoforms of TGF-b (TGF-b 1, -b 2, and -b 3) are expressed differentially at both the mRNA and protein levels during development of the heart (9). This suggests that these isoforms have different roles in regulating tissue development and growth. Therefore, Sharma and colleagues investigated distribution of the myostatin gene in other organ tissues using more sensitive detection techniques than that used by earlier researchers (8).
They found a DNA sequence in sheep and cow heart tissue that was identical to the respective skeletal muscle myostatin protein sequence, indicating the presence of myostatin gene in these tissues. In heart tissue from a Belgian Blue fetus, the myostatin gene deletion present in skeletal tissue was detected. They detected the unprocessed precursor and processed myostatin protein in normal sheep and cattle skeletal muscle, but not in that of the Belgian Blue. As well, only the unprocessed myostatin protein was found in adult heart tissue.
Animals with induced myocardial infarction (causing death of cells in heart tissue) displayed high levels of myostatin protein, even at 30 days postinfarct, in cells immediately surrounding the dead lesion. However, undamaged cells bordering the infarcted area contained very low levels of myostatin protein similar to control tissue. Considering the increase in other TGF-b levels in experimentally infarcted heart tissue (10), these growth factors may be involved in promotion of tissue healing.
Shaoquan and colleagues at Purdue University detected myostatin mRNA in the lactating mammary glands of pigs, possibly serving a regulatory role in the neonatal pig (12). They also detected similar mRNA is porcine skeletal tissue, but not in connective tissue. Most studies, in addition to this one, confirm that high levels of myostatin mRNA in prenatal animals and reduced levels postnatal at birth and postnatal reflect a regulatory role of myostatin in myoblast (muscle cell precursors) growth, differentiation and fusion.
A mutation in the myostatin gene in the two cattle breeds is not as advantageous as in mice. The cattle have only modest increases in muscle mass compared to the myostatin knockout mice (20-25% in the Belgian Blue and 200-300% in the null mice). Also, the cattle with myostatin mutations have reduced size of internal organs, reductions in female fertility, delay in sexual maturation, and lower viability of offspring (6). Although no heart abnormalities in myostatin-null mice were reported, the hearts in adult Belgian Blue cattle are smaller (11). Although the reduction in organ weight has been attributed to skeletal muscle mass increases, this has yet to be confirmed. Since there is evidence that the effects of myostatin mutation on heart tissue are variable in different species, there may be other possible tissue variabilities as well. Additionally, research detected myostatin mRNA in tissues other than skeletal muscle, demonstrating its expression is not relegated to skeletal muscle tissue as originally thought. Only further research will elucidate these possibilities.
Although several TGF-b superfamily members are found in skeletal and cardiac muscle tissue, their exact roles in development is not yet clear. Apparently, based on the early studies, the myostatin protein may have diverse roles in developmental and adult stage tissues. Sharma et al proposes that “myostatin has different functions at different stages of heart development” (8). As we shall see, the same can conceivably apply to skeletal muscle as well.
Myostatin and regulation of skeletal muscle
While many of the studies demonstrate that myostatin is involved with prenatal muscle growth, we know little of its association with muscle regeneration. Muscle regeneration of injured skeletal muscle tissue is a complex system and ability for regeneration changes during an animal’s lifetime. Exposure of tissues to various growth factors is altered during a lifetime. In embryos and young animals, hormones and growth factors favor muscle growth. However, many of these factors are downregulated in adults. Alteration in growth factors inside and outside of the muscle cells may diminish their capacity to maintain protein expression. Although protein mRNA may be detected within the cell, there are many sites of protein regulation beyond mRNA levels. As mentioned above, myostatin protein occurs in an unprocessed (inactive) and processed (active) form. Therefore, bioactivity of myostatin may be regulated at any point of its synthesis and secretion.
Keep in mind that nearly all regulatory systems in the body are under positive and negative control. This includes cardiac and skeletal muscle tissues. Myoblasts in developing animal embryos respond to different signals that control proliferation and cell migration. In contrast, differentiated muscle cells respond to another set of different signals. Distinct ratios of signals regulate the transition from undetermined cells to differentiated cells and ensure normal formation and differentiation in cellular tissues. However, many of the factors that regulate the various development pathways in muscle tissue are still poorly understood.
MyoD, IGF-I and myogenin (growth promoters in muscle cells) gene products are associated with muscle cell differentiation and activation of muscle-specific gene expression (14). Muscle-regulatory factor-4 (MRF-4) mRNA expression increases after birth and is the dominant factor in adult muscle. This growth factor is thought to play an important role in the maintenance of muscle cells. In addition to myostatin, there are other inhibitory gene products, such as Id (inhibitor of DNA binding). Although in vitro experiments are revealing the mechanisms of these specific proteins, we know less regarding their roles in vivo.
Although we know that lack of myostatin protein is associated with skeletal muscle hypertrophy in McPherron’s gene knockout mice and in double-muscled cattle, we know little about the physiological expression of myostatin in normal skeletal muscle. Recent studies in animal and human models indicate a paradox in myostatin’s role on growth of muscle tissue.
For example, evidence shows that myostatin may be fiber-type specific. Runt piglets, which have lower birth weights than their normal littermates, had lower proportions of Type I skeletal muscle fibers in specific muscles (12). Similar observations were made in rats where undetectable levels of myostatin mRNA in atrophied mice soleus (Type I fibers) (13). Transient upregulation of myostatin mRNA was detected in atrophied fast twitch muscles but not in slow twitch muscles. Thus, myostatin may modulate gene expression controlling muscle fiber type.
Studies also demonstrated lack of metabolic effects on myostatin expression in piglets and mice (12, 13). Food restriction in both piglets and mice did not affect myostatin mRNA levels in skeletal muscle. Neither dietary polyunsaturated fatty acids nor exogenous growth hormone administration in growing piglets altered myostatin expression (12). These and other studies strongly suggest that the physiological role of myostatin is mostly associated with prenatal muscle growth where myoblasts are proliferating, differentiating and fusing to form muscle fibers.
Although authors postulate that myostatin exerts its effect in an autocrine/paracrine fashion, serum myostatin has been detected demonstrating that it is also secreted into the circulation (8, 4). It is believed that the protein detected in human serum is of processed (active form) myostatin rather than the unprocessed form. High levels of this protein have been associated with muscle wasting in HIV-infected men compared to healthy normal men (4). However, this association does not necessarily verify that myostatin directly contributes to muscle wasting. We do not know if myostatin acts directly on muscle or on other regulatory systems that regulate muscle growth. Although several authors postulate that myostatin may present a larger role in muscle regeneration after injury, this has yet to be confirmed.
Myostatin and athletes
Further complicating the issue of myostatin’s role in regulation of muscle growth is the report by a team of scientists that mutations in the human myostatin gene had little impact on responses in muscle mass to strength training (15, unpublished data). Based on the report that muscle size is a heritable trait in humans (16), Ferrell and colleagues investigated the variations in the human myostatin gene sequence. They also examined the influence of myostatin variations in response of muscle mass to strength training.
Study subjects represented various ethnic groups and were classified by the degree of muscle mass increases they experienced after strength training. Included were competitive bodybuilders ranking in the top 10 world-wide and in lower ranks. Also included were football players, powerlifters and previously untrained subjects. Quadricep muscle volume of all subjects was measured by magnetic resonance imaging before and after nine weeks of heavy weight training of the knee extensors. Subjects were grouped and compared by degree of response and by ethnicity.
There were several genetic coding sequence variations detected in DNA samples from subjects. Two changes were detected in a single subject and another two were observed in two other individuals. They were heterozygous with the wild-type allele, meaning they had one allele with the mutation and the other allele was normal. The other variations were present in the general population of subjects and determined common. One of the variations was common in the group of mixed Caucasian and African-American subjects. However, the less frequent allele had a higher frequency in African-Americans. Although, as the authors comment, “these variable sites [in the gene sequence] have the potential to alter the function of the myostatin gene product and alter nutrient partitioning in individuals heterozygous for the variant allele”, the data from this and other studies so far show that this may not occur. This study did not demonstrate any significant response between genotypes and response to weight training. Nor were there any significant differences between African-American responders to strength training and non-responders or between Caucasian responders and non-responders.
Further research will be necessary to determine whether myostatin has an active role in muscle growth after birth and in adult tissues. To ascertain benefit to human health, we also need to discover its role in muscle atrophy and regeneration after injury. Only extended research will reveal any such benefits.
The future of myostatin
Now that we have reviewed some of the biology of the myostatin protein, its gene, and the relevant scientific literature, what are the implications for its application?
Many authors of the myostatin studies have speculated that interfering with the activity of myostatin in humans may reverse muscle wasting disease associated with muscular dystrophy, AIDS and cancer. Some predict that manipulation of this gene could produce heavily muscled food animals. Indeed, current research is underway to investigate and develop these potentialities. Sure enough, a large pharmaceutical company has recently applied for a patent on an antibody vaccination for the myostatin protein.
A medical doctor and author of weight training articles asserts that overexpression of myostatin is to blame for weight lifters that have trouble gaining muscle mass. The spokesperson for a supplement and testing lab erroneously implied that the “rarest” form of mutation in the myostatin gene is responsible for a top competitive bodybuilder’s massive muscle gains, not taking into account the performance-enhancement substances the bodybuilder may be using. The public media has, of course, predicted that “steroid-popping” athletes will take advantage of myostatin inhibitors to gain competitive edge (3).
Many of these assertions are unfounded or they misrepresent the science. Granted, the possibility exists that manipulation of the myostatin gene in humans may be a key to reversing muscle-wasting conditions. However, too little is still yet unknown regarding myostatin’s role in muscle growth regulation. It is imperative that research demonstrates that the loss of myostatin activity in adults can cause muscle tissue growth. Likewise, research must also prove that overexpression or administration of myostatin causes loss of muscle mass. Also important is to know if manipulation of myostatin will interfere with other growth systems, especially in other tissues, and result in abnormal pathologies. Although McPherron’s gene knockout mice did not experience any other gross abnormalities, mice are not humans.
We do not fully understand the roles of myostatin in exercise-induced muscle hypertrophy or regeneration following muscle injury. Until we do, it may be premature to blame the lack of hypertrophy in weightlifters on overexpression of myostatin. Nor does the research support the claim that a top bodybuilder’s muscle mass gains are resultant of a detected mutation in the myostatin gene. The research simply does not advocate blaming genetic myostatin variations as a source of significant differences in human phenotypes.
Considering the history of the athlete’s propensity, in the public eye, to abuse performance-enhancement substances, the media’s prediction of myostatin-inhibitor may or may not be warranted. We all know that today’s athletic arena demands gaining the competitive edge to maintain top level competition. For many athletes, that is accomplished by supplementing hard training with substances that enhance growth or performance. Whether or not myostatin inhibitors will be added to the arsenal of substances is difficult to predict. Until science reveals the full nature of this growth factor and its role in the complex regulation of muscle tissue, and researchers determine its therapeutic implications, we can only surmise. Despite attempts to tightly control any pharmaceutical uses of myostatin protein manipulation, they will likely surface at some point in the black market world of bodybuilding supplements. Let us hope that science has determined the side effects and the benefits by that point.
References
McPherron, AC, AM Lawler, SJ Lee. Regulation of skeletal muscle mass in mice by a new TGF-b superfamily member. Nature 1997, 387:83.
McPherron, AC, SJ Lee. Double muscling in cattle due to mutations in the myostatin gene. Proc Natl Acad Sci USA 1997, 94:12457
John Hopkins Magazine, June 1997. URL: http://www.jhu.edu./~jhumag/0697/web/science.html
Gonzalez-Cadavid, NF, WE Taylor, K Yarasheski, et al. Organization of the human myostatin gene and expression in healthy and HIV-infected men with muscle wasting. Proc Natl Acad Sci 1998, 95:14938.
Grobet, L, LJR Martin, D Poncelet, et al. A deletion in the bovine myostatin gene causes the double-muscled phenotype in cattle. Nature Genet 1997, 17:71.
Menissier, F. In: Muscle Hypertrophy of Genetic Origin and its Use to Improve Beef Production, eds. King, JWB and F Mennissier. Nijhoff, The Haugue, The Netherlands, pp. 23-53.
Kambadur R, M Sharma, TPL Smith, JJ Bass. Mutations in myostatin (GDF8) in double-muscled Belgian Bllue and Piedmontese cattle. Genome Res 1997, 7:910.
Sharma M, R Kambadur, KG Matthews, et al. Myostatin, a transforming growth factor-b superfamily member, is expressed in heart muscle and is upregulated in cardiomycetes after infarct. J Cell Physiol 1999, 180:1.
Millan FA, F Denhes, P Kondaiah, et al. Embryonic gene expression pattern of TGF-b 1, b 2, and b 3 suggest different developmental function in vivo. Development 1991, 111:131.
Sharma, HS, M Wunsch, T Brand, et al. Molecular biology of the coronary vascular and myocardial responses to ischemia. J Cardiovas Pharmacol 1992, 20:S23.
Bocard R. 1981. Facts and reflections on muscular hypertrophy in cattle: double muscling or culard. In: Developments in Meat Science, Vol. 2. Lawrie R, ed. Applied Science Publishers, London, pp. 1-28.
Shaoquan, J, RL Losinski, SG Cornelius, et al. Myostatin expression in porcine tissues: tissue specificity and developmental and postnatal regulation. Am J Physiol 1998, 275:R1265.
Carlson, JC, FW Booth, SE Gordon. Skeletal muscle myostatin mRNA expression is fiber-type specific and increases during hindlimb unloading. Am J Physiol 1999, 277:R601.
Marsh, DR, DS Criswell, JA Carson, FW Booth. Myogenic regulatory factors during regeneration of skeletal muscle in young, adult and old rats. J Appl Physiol 1997, 83:1270.
Ferrell, RE, V Conte, EC Lawrence, et al. Frequent sequence variation in the human myostatin (GDF8) gene as a marker for analysis of muscle related phenotypes. Genomics, in press.
Loos, R, M Thomis, HH Maes, et al. Gender-specific regional changes in genetic structure of muscularity in early adolescence. J Appl Physiol 1997, 82:1602.
About the author
Elzi has spent the last several decades trying to determine where 'home' is: from New York, Maine, California, Oregon and now Texas. As well, her career has encompassed tool & die apprentice, forest ranger, assistant extension agent, mother, sheep and horse rancher, and mad research scientist. She has also been a competitive bodybuilder, but has found true joy in powerlifting. When Elzi is not playing fetch with her 1200 lb four-footed buddy, she is most happy in the gym and in a research lab.
Leave a Reply
You must be logged in to post a comment.